
Biochemistry 200
January 19-22, 1999
Julie Theriot
Carbohydrate and Energy Metabolism
Recommended reading:
Stryer (4th edition) or Mathews and van Holde
Carbohydrates: Chapter 18, pp. 463-474 Chapter 8, pp. 260-281
Glycolysis: Chapter 19, pp. 483-491 Chapter 13, pp. 433-450
Regulation of glycolysis: Chapter 19, pp. 493-495 Chapter 13, pp. 450-453
Glycogen: Chapter 23, pp. 581-589 Chapter 13, pp. 458-465
and Chapter 16, pp. 550-556
Hormones and cAMP: Chapter 23, pp. 590-599 Chapter 23, pp. 780-790 and 797-801
Questions? After class and in the Beckman Center, Room 473A. Phone 725-7968.
Introduction to metabolism
Metabolism is the integrated and purposeful set of chemical reactions that enables us to change the food we consume into the substances that make us up. Energy to maintain life is derived from the chemical bond energy of food.
An organic substance like sugar can be burned, oxidized, to release this chemical bond energy all at once in the form of heat. Since we live in an oxygen-rich world, oxidation proceeds spontaneously. Thus the more reduced form of a molecule, the more electron-rich form, is considered to be at a higher energy state.

In living cells, sugar is oxidized in a stepwise process. Enzyme-catalyzed reactions ensure that a large part of the free energy released by oxidation of food is captured in a chemically useful form. This is done by coupled reactions, in which an energetically favorable reaction (such as oxidation of glucose) is coupled to an energetically unfavorable one that produces an activated carrier molecule. The activated carrier molecules store energy in an easily exchangeable form, either as a readily transferable chemical group (such as the
g-phosphate on ATP) or as high-energy electrons (as on NADH).ATP (adenosine triphosphate) NAD+ (nicotinamide adenine dinucleotide) NADH

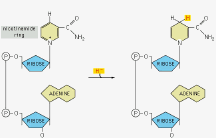
ADP
Although innumerable types of organic molecules can be degraded and synthesized by cells, most interchanges of activated groups in metabolism are accomplished by a rather small number of carrier molecules. For example, proteins, carbohydrates, and fats are all broken down to generate ATP, NADH, and acetyl CoA.

In metabolic pathways, the product of one reaction becomes the substrate for the next reaction. Different metabolic pathways are interconnected, and flux of intermediates through them is regulated by nutrient availability and hormone signals. Under different conditions, the same molecules might be either degraded or synthesized, depending on the body’s immediate needs. For example, all cells break down glucose to pyruvate to generate energy, a pathway called glycolysis. However, if a person has not eaten and blood glucose is low, the cells of the liver will use energy to synthesize glucose from pyruvate, a pathway called gluconeogenesis, and deliver it into the blood. This ensures that the brain will always have an adequate energy supply, at the expense of other organs if necessary. Degradation and synthesis generally take place by distinct pathways.
Carbohydrates
This set of lectures will describe the metabolism of carbohydrates. Metabolism of fats and proteins will be covered later in the course. Carbohydrates have the general formula (CH2O)n, carbon + water. Monosaccharides (simple sugars) can have 3 to 8 carbons, and contain multiple hydroxyl groups as well as either an aldehyde group (aldoses) or a ketone group (ketoses).


glucose fructose
(6-carbon aldose) (6-carbon ketose)
In aqueous solution, one of the hydroxyl groups tends to react with the aldehyde or ketone group, closing the linear molecule into a ring. In an equilibrium mixture, only about 1% of the sugar exists in the open chain form, but the two forms interconvert rapidly.

In the closed ring form, the hydroxyl group on the carbon that carries the aldehyde or ketone can lie either below the plane of the ring or above it. The two positions are called
a and b. They also rapidly interconvert, but once one sugar is linked to another, the a or b form is frozen. 
The
a or b hydroxyl group can react with any hydroxyl group on a second sugar molecule to form a glycosidic bond, releasing water. Sucrose is a disaccharide formed by an a linkage between carbon 1 of glucose and carbon 2 of fructose. Lactose is formed by a b linkage between carbon 1 of galactose and carbon 4 of glucose. When the hydroxyl group on a carbon that carries a carbonyl is available for such a reaction, the sugar is said to be "reducing". Sucrose is a nonreducing sugar. 
Oligosaccharides and polysaccharides are formed from multiple sugars linked in a chain by a series of glycosidic bonds. The chemical nature of a polysaccharide is determined by its constituent subunits and the types of linkages between them. For example, a chain of glucose molecules held together by
b 1g4 glycosidic bonds is cellulose, a major component of plant cell walls. A chain of glucose subunits held together by a 1g4 glycosidic bonds is a-amylose, or starch. 
When carbohydrate-containing food is consumed, polysaccharides and disaccharides are hydrolyzed to their component monosaccharide subunits by digestive enzymes in the saliva in the lumen of the small intestine. The enzymes are stereospecific.
a-amylase in the saliva and secreted by the pancreas into the intestine will hydrolyze a 1g4 bonds but not b 1g4 bonds, therefore we can digest starch but not cellulose.A variety of specific enzymes on the surface of the intestinal cells hydrolyze various disaccharides. Lactose in milk is a major source of nutrition for young mammals including humans, and it is split by
b-galactosidase (also called lactase) in the intestine into galactose and glucose. Most adult mammals, including the majority of adult humans, stop secreting this enzyme, and become lactose intolerant. Undigested lactose is osmotically active, drawing water into the large intestine and causing diarrhea. Lactose is readily fermented by the bacteria living in the large intestine, generating CO2 and H2 gas, resulting in bloating and flatulence.Two types of glucose carriers enable gut epithelial cells to transfer glucose and other monosaccharides across the gut lining into the bloodstream. Glucose is actively transported into the cell by Na+-driven cotransporters at the apical surface, a process that requires energy (in the form of the Na+ gradient) since glucose is moving from a region where its concentration is low, in the lumen, to a region where its concentration is high, in the intestinal cell cytoplasm. Glucose is released by the cell into the bloodstream by passive transport down its concentration gradient, mediated by a different glucose transporter. Most other cells take up glucose from the bloodstream via passive transport, since their cytoplasmic glucose concentration is lower than the glucose concentration in blood.

Glycolysis
All tissues in the body break down glucose to provide energy and intermediates for other metabolic and biosynthetic pathways. Virtually all sugars can be converted to glucose, so the process of glycolysis is central to carbohydrate metabolism. For cells that lack mitochondria, such as red blood cells, glycolysis is the only available means for generating ATP. Consequently, people with defects in the enzymes that catalyze glycolysis frequently suffer from hemolytic anemia.
During glycolysis, the six-carbon sugar is split into two molecules of pyruvate, which each contain three carbons. Glycolysis proceeds in ten steps. Two molecules of ATP are hydrolyzed to provide energy to drive the early steps, but four molecules of ATP (as well as two molecules of NADH) are produced in the later steps.

Step 1: Phosphorylation of glucose.
Glucose is phosphorylated by ATP to form a sugar phosphate. Phosphorylated glucose is not recognized by glucose transporters, so this step effectively traps glucose inside the cell. In most cells, this reaction is catalyzed by the enzyme hexokinase. In liver cells and in the
b cells of the pancreas, a different enzyme is used, glucokinase. The different properties of these enzymes are important for the proper regulation of carbohydrate metabolism (which we will discuss below). This is the first energy-consuming step of glycolysis. The rate of entry of glucose into the glycolytic pathway can be regulated at this step. 
In these diagrams the symbol ![]() represents PO32-.
represents PO32-.
The binding of glucose to hexokinase causes a large conformational change in the enzyme, closing the two halves of the protein together like a clamp.

This conformational change creates a binding site for ATP, and effectively excludes water from the active site. Transfer of the phosphate group from ATP to glucose also requires the presence of magnesium.

Step 2: Isomerization of glucose-6-phosphate to fructose-6-phosphate
A readily reversible rearrangement moves the carbonyl oxygen from carbon 1 to carbon 2, transforming the aldose into a ketose.

The enzyme stabilizes the ene-diol intermediate, using general acid-base catalysis. The catalytic residues are lysine (BH+) and glutamate (B’).

Step 3: Phosphorylation of fructose-6-phosphate
The new hydroxyl group on carbon 1 is phosphorylated by ATP. This is the second energy-consuming step, and is the rate-limiting step in glycolysis. Regulation of the enzyme responsible, phosphofructokinase, is the most important control point. The reaction is similar to the hexokinase reaction in step 1, and also requires Mg2+.

Step 4: Cleavage of fructose-1,6-bisphosphate
Aldolase catalyzes the reversible cleavage of the six-carbon sugar into two 3-carbon species, dihydroxyacetone phosphate (DHAP) and glyceraldehyde-3-phosphate (G3P). This is the only step in glycolysis where a carbon-carbon bond is split. The product G3P can proceed directly through the next steps of glycolysis, but DHAP cannot.

Aldolase forms a covalent bond with its substrate, forming a Schiff base.

Step 5: Isomerization of dihydroxyacetone phosphate
DHAP is converted into G3P by triose phosphate isomerase; that is, the ketose is isomerized to the equivalent aldose. This reaction is analogous to step 2, and is also readily reversible.

Triose phosphate isomerase (TIM) stabilizes the ene-diol intermediate, using concerted general acid-base catalysis. TIM is one of the most efficient enzymes known, approaching "kinetic perfection" (the rate of the reaction is limited by the diffusion rate of the substrate).

Step 6: Oxidation and phosphorylation of glyceraldehyde-3-phosphate
The two molecules of G3P are oxidized. This is the first part of the energy-generating phase of glycolysis. NAD+ accepts two electrons and one proton to become NADH, and a new high-energy anhydride linkage to phosphate is created.

Glyceraldehyde 3-phosphate dehydrogenase forms a covalent bond to the substrate through a reactive -SH group on the enzyme, and catalyzes its oxidation while still attached. The reactive enzyme-substrate bond is then displaced by an inorganic phosphate ion to produce a high-energy phosphate intermediate, 1,3-BPG.

Step 7: Transfer of high-energy phosphate from 1,3-bisphosphoglycerate to ADP
This step generates two molecules of ATP per molecule of glucose, (one molecule of ATP per molecule of 1,3-BPG). At this point, the ATP’s that were "invested" in steps 1 and 3 have been recouped. Combined, step 6 and step 7 oxidize an aldehyde to a carboxylic acid, releasing energy that is stored in the form of NADH and ATP.

Arsenate (AsO43-) closely resembles inorganic phosphate and can substitute for it in step 6. The product of this reaction, 1-arseno-3-phosphoglycerate, is unstable and decomposes in water to generate 3-phosphoglycerate, without generating an ATP. Thus although the glyceraldehyde-3-phosphate is oxidized, oxidation and phosphorylation reactions are uncoupled, and arsenic is a potent cellular poison.
Step 8: Shift of the phosphate group from carbon 3 to carbon 2

This freely reversible reaction isomerizes 3-phosphoglycerate to 2-phosphoglycerate. The enzyme phosphoglycerate mutase has a phosphoryl group attached to a histidine residue at its active site, which it transfers to form the obligate intermediate 2,3-bisphosphoglycerate. The phosphoryl group is therefore not transferred directly from carbon 3 to carbon 2 within a single substrate molecule; rather, the phosphoryl group of the incoming molecule of 3-phosphoglycerate ends up on carbon 2 of the next substrate whose conversion is catalyzed by the enzyme.

Occasionally, 2,3-BPG dissociates from the enzyme, leaving it in an inactive form. For this reason, the cell needs to maintain trace levels of 2,3-BPG to regenerate the phosphoenzyme by the reverse reaction. 2,3-BPG is synthesized for this purpose by a separate pathway, involving isomerization of 1,3-BPG by bisphosphoglycerate mutase. This reaction is particularly important in red blood cells, where 2,3-BPG has a role in regulating the affinity of hemoglobin for oxygen.
Step 9: Dehydration of 2-phosphoglycerate
The removal of water from 2-phosphoglycerate changes the relatively low-energy phosphate ester linkage to a high-energy enol phosphate linkage. This reaction is readily reversible.


Step 10: Formation of pyruvate
In the last step of glycolysis, the high-energy enol phosphate is transferred to ADP, generating another molecule of ATP (two molecules of ATP per original molecule of glucose). This step is effectively irreversible, and pyruvate kinase is the third regulated enzyme of glycolysis.

Fates of pyruvate
In most animal and plant cells, glycolysis is a prelude to the final stage of energy production, which occurs in the mitochondria. Pyruvate is imported into mitochondria, where it is converted into acetyl-CoA. Acetyl-CoA is an important building block in biosynthetic reactions. It is also a major fuel for the TCA cycle. Within the mitochodria, acetyl-CoA is completely oxidized by the TCA cycle and oxidative phosphorylation into CO2 and water, generating large amounts of ATP. Reducing equivalents present in the cytoplasm as NADH (generated in step 6) can be shuttled into the mitochondria by two different mechanisms that regenerate NAD+, and the resulting reducing equivalents in the mitochondria can be used directly in oxidative phosphorylation to generate ATP. These pathways require molecular oxygen (O2), and will be detailed in the next set of lectures on aerobic metabolism.
In human cells that lack mitochondria (such as red blood cells and the lens cells in the eye), or when there is little or no molecular oxygen available (such as in skeletal muscle during vigorous exercise), pyruvate remains in the cytoplasm. Under these conditions, glycolysis can be limited by the amount of NAD+ available for the oxidation of glyceraldehyde-3-phosphate in step 6. NADH generated in this step can be to reduce pyruvate to lactate, a reaction catalyzed by lactate dehydrogenase, regenerating NAD+.

Lactic acid can accumulate in muscle, causing cramps. Most lactate eventually diffuses into the bloodstream. In the liver, lactate can be reconverted to glucose by the process of gluconeogenesis, which will be described later. Lactate is reoxidized to pyruvate in heart muscle, and used to make ATP in the TCA cycle. Excessive lactate production can result in lactic acidosis, where blood pH drops significantly. High blood lactate can be a sign of general tissue hypoxia, such as during shock or pulmonary failure.
In yeast and some bacteria, pyruvate is converted first into acetaldehye by pyruvate decarboxylase, and then reduced to ethanol by NADH. Like the production of lactate, the conversion of glucose to ethanol involves no net oxidation and can occur in the absence of O2.
Summary of glycolysis
